
为了对抗病毒,人类免疫系统都进化出哪些机制?
录入者:15959541500 人气指数: 次 发布时间:2020年11月12日

出品:新浪科技《科学大家》、未来论坛
主讲嘉宾:祈海 清华大学终身教授
不管在美国还是在中国,每个人每天不可避免地听到新冠病毒相关新闻,不论是做科研还是不做科研,大家都好奇疫情到底什么时候能结束。不看历史不可知未来,历史上人类有过很多次大的瘟疫。
从免疫学工作者的角度,我发现其中一个很明显特点:在文艺复兴时期,欧洲所流行的鼠疫据不完全统计导致将近三分之一的欧洲人口死亡。佛罗伦萨的黑死病,尽管当时死了十几万人,这个瘟疫还是过去了,那个时候显然没有现在的免疫学知识。实际上公共卫生的改善,城市的治理使得人和鼠能够分开,生活用水得到更洁净地处理,这些公共卫生的改变使得很多历史上重大的瘟疫,即使没有今天科学知识也能得到控制。这一点是值得任何人都关注的,即使没有治疗方法或者疫苗开发,也并不是说不能控制这个疾病。
如果从达尔文的眼里看,病毒的肆虐一定是被想象成一种种属之间物竞天择的斗争。下图是我2015年到肯尼亚草原上看动物有幸拍摄到猎豹捕猎的过程。

这是物竞天择的物种间的斗争,今天疾病的流行当然是我们和病毒之间的斗争。千百万年来演化出来的免疫系统到底有哪些特点?如何和病毒做斗争?我们是不是可以理解成,我们得病是免疫系统不够强大,不足以抵抗这种新冠?另外主持人颜宁老师提到,即使是学生物的朋友,当面对免疫系统的时候还是听起来非常复杂,这背后有什么样的生物学逻辑,来支撑免疫系统与病毒做斗争呢?当然研究免疫系统,最终是希望能有更好的方法,通过疫苗以及它的方式,能够帮助免疫系统更强地与病毒做斗争。
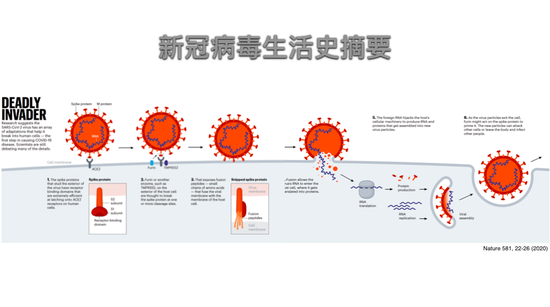
讲免疫学之前,我们先看一下新冠病毒到底是什么样?之所以叫它冠状病毒,是因为它在电镜下看起来像一个花冠,它的Spike蛋白长的也像花冠。它需要和宿主的受体转换酶2进行结合才能够进入细胞。我们知道病毒自己不可以独立生存,它需要到宿主细胞里面,利用宿主细胞的生物学的机器来复制自己。
这次新冠病毒有一个非常特殊的地方:跟其它冠状病毒相比,它的Spike 蛋白上多出来一个酶切位点,使它和受体结合以后,产生融合构象的效率非常高,所以它的感染性也很高。病毒颗粒和宿主细胞融合就可以把它的遗传物质注入到宿主细胞里。
新冠病毒是一种正链的RNA病毒,所以它的遗传物直接可以作为模板进行翻译,产生病毒的蛋白。病毒遗传物质注入到宿主细胞以后翻译成病毒的蛋白,并且遗传物质被复制,进一步可以组装成新的病毒颗粒从宿主细胞里释放。宿主细胞本来应该正常工作,现在被绑架来生产病毒,自然就会产生病理反应,机体功能也会发生改变。所以什么样的细胞能够被病毒感染,很大程度上决定了病毒会导致哪些器官组织出现问题。
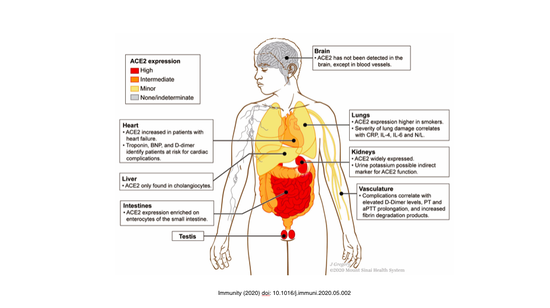
上图是ACE2表达谱,可以看到除了肺细胞,肾脏、肠道、肝以及心血管系统的内皮细胞,都表达ACE2,包括男性生殖腺。所以ACE2的广泛表达使得潜在被感染的细胞很多,这样可以理解病毒造成多器官病变的病理学机理。病毒感染以后,实际上每个细胞本身还是有固有的免疫的机制的,可以感知病毒。
免疫系统的模式识别:专门感知病原体
在固有免疫系统里面有个非常重要的概念是模式识别,机体里的每一个细胞都有一套机器,专门用来感知病原体入侵。不管是新冠病毒还是哪一种病毒,每个细胞都继承了一套模式识别系统,使得细胞可以知道是不是有病原入侵。具体到RNA病毒,至少有四种分子机器。对于不是学生物的人来说,我们知道这个概念就好。比如有RIG-I、MDA5等。这些分子机器试图检测细胞具备哪些病毒特点。RNA病毒为了复制,一定会形成RNA的双链结构,正常情况下我们的细胞里很少有长的RNA双链结构,这样的结构就会导致这些分子机器和它进行结合形成多聚体,这些多聚体有酶学活性,产生了下游信号传导过程,使得被感染细胞本身出现了固有抗病毒特性,试图抑制病毒复制。
还有各种各样的机制,其中很重要的一点,新冠病毒进入细胞以后,RIG-I或者MDA5活化以后就会导致细胞产生I型干扰素,这是一个非常重要的抗病毒途径。I型干扰素的受体几乎在所有的细胞上都并表达,也就是任何一个细胞都可以接受I型干扰素的信号,从而进入一种高度戒备的抗病毒状态。I型干扰素的信号转导下游有很多抗病毒基因,比如说有一些是A酶,能够直接切断那些长链的RNA,自然就不能进一步作为模板来复制,病毒复制就会受到抑制。
I型干扰素从任何一个细胞产生,可以作用在它旁边的细胞,一个细胞如果被感染了,它产生这种抗病毒机制,就会使旁边的细胞都处于更高的警戒状态,增强抗病毒能力。所以细胞有模式识别系统使得它本身产生了固有抗病毒机制。在最开始与病毒搏斗。当然这显然不够。病毒复制的速度非常快,那么这个过程中被感染的细胞也通过像I型干扰素这样的炎性分子招募其它的更经典意义上的免疫细胞,产生所谓的炎症。
炎症到底是什么?
炎症一词大家既熟悉又陌生,因为它会在所有疾病的情况里面都会出现,那么炎症到底是什么?
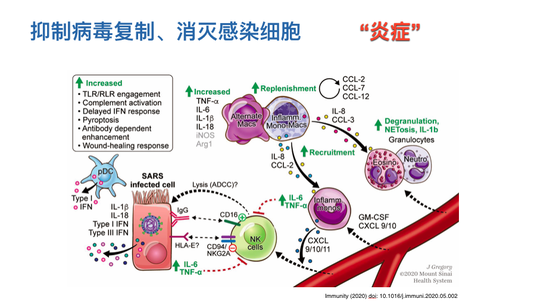
在病毒免疫反应里面,炎症是那些参与控制病毒、抑制病毒复制、杀灭病毒感染的免疫细胞在组织里浸润的过程。中性粒细胞有存脂功能,也可以产生很多细胞因子、趋化因子,招募更多的细胞。还有自然杀伤细胞,它可以直接杀伤潜在感染的细胞。如果把感染的细胞杀掉,当然可以控制感染的过程。大家也注意到各种各样的黏性细胞,巨噬细胞、中性粒细胞。
这些免疫细胞虽然有不同的功能,但可以发现它们有非常相似的特点,都在产生各种各样的炎性因子和趋化因子,它们互相之间会招募更多同类的细胞,同时他们在进行抗病毒或者是阻止炎症反应过程中会帮助其它细胞,所以它们有一个协同作用,使得那些被病毒感染的细胞被包裹在这样一个炎症的环境,病毒复制过程可以被一定程度地抑制。
如果我们考虑病毒扩增是指数增长,一个细胞可以产生成千上万个病毒颗粒,成千上万个病毒颗粒可以再感染更多的细胞,然后滚雪球一样的指数增长。如果考虑固有抗病毒机制,也就是某一个被感染的细胞产生I型干扰素,实际上是线性的过程。一个细胞顶多把自己病毒的复制抑制住,它产生的I型干扰素影响周围的细胞,但是这是线性的倍增。炎症反应招募了很多细胞,它们之间有协同作用,是乘积关系,比如中性粒细胞和嗜酸性粒细胞之间有一个协同作用,但是追不上指数的病毒的扩增。
如果在病毒复制非常早期地情况下,即病毒还没有开始复制时候就产生了很强的炎症反应,可能病毒就会被抑制,否则病毒的指数扩增会击穿线性或者乘积性的保护机制。因此接下来抗原特异的免疫反应很重要。
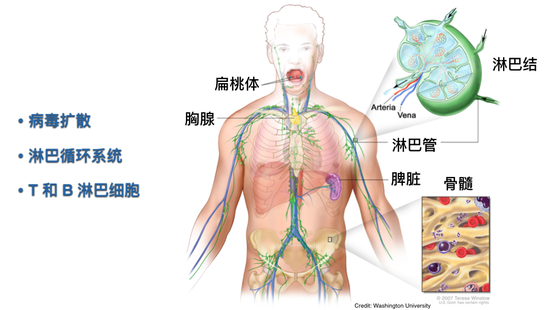
新冠病毒在肺里开始的指数扩增击穿了细胞固有的抗病毒机制,也不能被炎症细胞浸润给控制住,它向哪儿跑呢?
从肺里面出现的病毒,它主要的扩散的途径是通过两个循环细胞,血液循环细胞和淋巴循环细胞。在生物进化过程中,体积比较大的动物用淋巴循环细胞来收集组织间液,维持水循环平衡。它也被获得性免疫系统用来收集抗原。在图中可以看到,淋巴循环的淋巴管分布在各种各样的组织里,包括肺组织。它通过右心的负压使得组织间液被抽进淋巴循环,然后组织间液就会通过淋巴管流经淋巴结,淋巴结里有很多淋巴细胞,可以识别潜在的抗原。
脾脏是更大的二级淋巴器官,它是通过血液把里面潜在的病原捕获住,然后让获得性免疫系统可以识别。淋巴细胞大致分为T淋巴细胞和B淋巴细胞。它们从哪儿来?最开始都是从骨髓的造血干细胞来,B细胞从骨髓里分化发育,在脾脏里成熟。T细胞最开始的前体细胞也是从骨髓来,但是它在胸腺里进行成熟和发育,而后通过血液循环进入淋巴循环,在没有抗原或者病毒来的情况下,它就在血液循环和淋巴循环这两个循环里周而复始,试图寻找潜在可能识别的抗原。
如果病毒来了,有一些病毒会进入脾脏或者淋巴结,那么这些细胞就可以识别它。所以激活淋巴细胞是固有的免疫机制以及炎症反应之后非常重要的一个事件。
淋巴结里的免疫反应是如何发生的?
树突状细胞是一类抗原提呈细胞,它分布在不同组织里,比如肺和支气管的上皮细胞下面就分布着这样的细胞,如果病毒来了树突状细胞可以捕获病毒本身或者被病毒感染的细胞,从而把病毒的抗原信息捕捉住。这样的细胞在被病毒激活或者炎症反应激活以后会顺着淋巴管迁徙到淋巴结里面去。刚才讲了病毒本身如果复制扩散也可以通过淋巴管直接进入淋巴结,淋巴结里也有树突状细胞,它也可以捕获这些病毒,从而把这些抗原信息提呈出来。树突状细胞捕获了这些病毒所特有的某些信息,然后把信息传递给T淋巴细胞,T淋巴细胞可以对这样的信息产生反应。我们身体里有成千上万不同种的T淋巴细胞,每个细胞只有一种抗原受体,可以识别一种抗原。
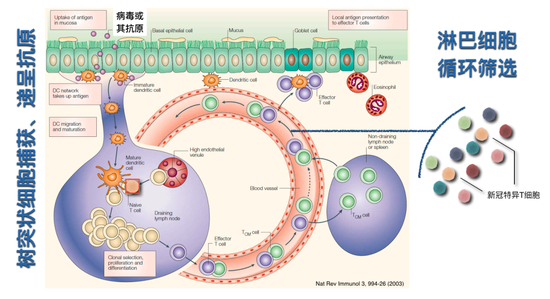
上图画的两种肉色细胞,我们就叫它新冠病毒特异T细胞,它是一开始和识别其它抗原的细胞混在一起,免疫反应的发生是让树突状细胞把新冠特异T细胞筛选出来,让它特异活化并参与免疫反应。比如新冠病毒感染了,可以识别新冠病毒的T细胞流经肺部淋巴结,在这里撞上了在呈递新冠病毒抗原的树突状细胞,这时候它就可以被树突状细胞上面呈递的新冠病毒特殊的抗原信息所激活。
白颜色的树突状细胞被表征成带有新冠病毒的树突状细胞。绿颜色的细胞就是刚才已经被循环的筛选到的可以特异识别新冠病毒的T淋巴细胞。这时候会发生什么呢?这个细胞和抗原呈递的树突状细胞会发生很重要的事件,树突状细胞会用主要组织相容性抗原分子MHC(抗原呈递的分子)(如图)把新冠病毒的抗原肽呈递给绿颜色细胞。形成T细胞要进行识别的分子复合物。
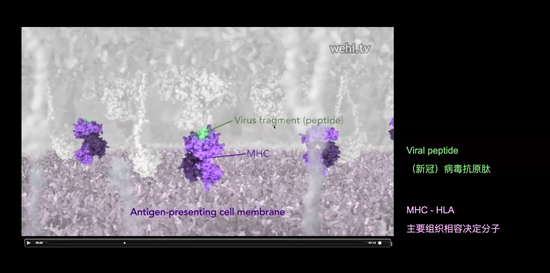
每一个T细胞都会有一种T细胞受体。什么意思呢?我们叫它克隆。每一个T细胞只有一种T细胞抗原受体,它可以识别一种抗原肽和MHC的组合,也就是识别新冠抗原肽和MHC组合的T细胞受体,不能去识别HIV的抗原肽和MHC组合。一个T细胞有一个受体,T细胞受体能够识别MHC和病毒的抗原肽然后被激活。
当然这个激活过程还需要很多其它辅助受体,比如具体情况下我们讲细胞毒性T细胞,它通过CD8杀伤细胞辅助受体,结合到MHC上面,这时候产生了很强的激活信号,这个信号就告诉T细胞应该激活了,而一个抗原呈递细胞表面可能有很多个抗原肽和MHC的复合物,很多个T细胞受体就会被这个过程激活,形成聚集物,产生很强的激活信号, T细胞就会开始要细胞增殖。这个增殖我们叫克隆增殖,也就是同样的细胞复制它自己,原来只有一个可以识别病毒抗原的细胞,现在变成很多个,呈指数扩增。
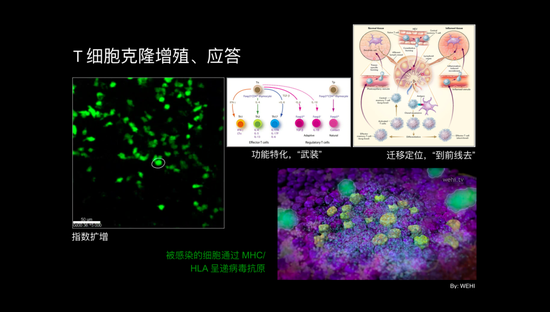
上图是真实实验动物体内拍的免疫反应中T细胞进行的克隆增殖,可以看到这个过程非常快。事实上在成年动物的体内几乎没有任何其他地方,可以看到这么迅速和大量的细胞增殖。这就是第一个免疫反应过程中产生了指数增长的效应的过程。
增殖过程中T淋巴细胞还要进行功能特化,换句话说它得把自己“武装”起来,它还得能够控制病毒。它在一个淋巴结里发生,要跑到肺里去才能控制最开始病毒感染的地方,它去了前线以后怎么控制病毒的过程呢?用这样一个模式显示,不同颜色的细胞都是被感染的细胞,已经被活化起来的T细胞到这儿来要杀伤被病毒感染的细胞。
怎么知道谁是被病毒感染细胞的呢?这个过程和它一开始被树突状细胞活化的生物学过程是一样的。感染的细胞表面也要表达MHC分子。因为这是一个病毒感染细胞,病毒细胞的胞浆里当然有病毒制造的病毒蛋白,它被MHC分子提呈出细胞表面,T细胞过去被树突状细胞活化时就能识别到病毒肽和MHC的复合物。
所以在这里只有感染的细胞才可能会有这样的呈递,原则上活化的T细胞只会杀掉这种被感染的细胞,从而实现一种特异地清除被感染细胞的杀伤效应。这个过程使得T细胞可以特异识别被感染的细胞,然后特异地向被感染细胞传递杀伤颗粒,从而把这个细胞杀掉。通过这样的过程我们可以看到,获得性免疫反应通过淋巴细胞应答,提供了指数增长效应。它和固有的抗病毒机制,再加上炎症反应放大结合在一起,使得我们可以追赶在体内进行指数扩增的病毒。
理想情况下,这个过程使得病毒受到抑制,病毒载量会降低,被感染细胞变少,机体开始恢复。如果一切顺利的话,固有抗病毒机制就会产生,I型干扰素分泌减少,炎症反应会消退,同时我们的组织开始修复。病毒被清除过程中,它自己也有程序化死亡的过程,这些细胞由于没有了病毒抗原就逐渐地消退掉,最终形成记忆T细胞,这是理想的情况,一次完整的免疫反应是:我们被感染,然后产生了获得性免疫,指数增长,杀死并清除病毒,最后形成免疫记忆系统,下一次再感染的时候就可以很快地抑制住。
当然这是理想情况。病毒复制很快,可以通过随机突变去寻找、抵抗免疫效应及抑制的过程,我们姑且叫它病毒抗T细胞免疫。病毒有很多手段抵抗免疫过程,很重要的一部分是从抗原识别角度来的。T细胞为了活化,需要用T细胞受体识别感染的细胞或者细胞上呈递的病毒编码抗原肽以及抗原呈递的MHC分子。
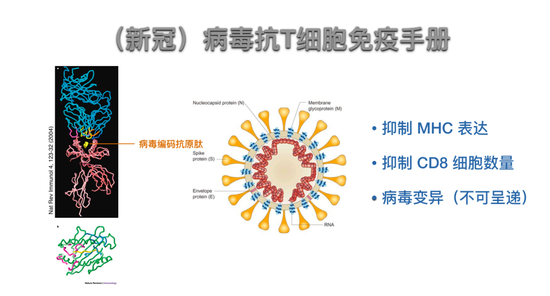
可以想像,任何一个MHC分子所呈递的病毒的抗原肽都是有限的。抗原肽都是由基因编码的,病毒进行突变可以非常快地把任何一个MHC分子进行呈递的抗原肽突变掉,这样病毒可以没有能呈递的抗原肽,也就是CD8细胞不会被活化了,这是非常可怕的。另外,所有病毒都能抑制被感染的细胞表达MHC,这也是试图拮抗T细胞免疫的手段之一。病毒还可以产生很多效应,使得机体的细胞数量变少。当然最可怕的是病毒变异使得可以被呈递的抗原肽变得不可被呈递。
免疫系统如何更好地拮抗病毒?
病毒和免疫系统既然是矛和盾之间的关系,那免疫系统在演化的过程当中有什么样的机制能够拮抗病毒的特性呢?这就涉及到MHC的同种异体多样性。并不是每个人都只有一个MHC,不同人群当中可以看到各种不同的MHC分子。
通过东亚人群和象牙海岸的人不同的MHC分子的频数,我们可以看到B52,在东亚人群里大概20%,5个人里有1个是有B52,但是象牙海岸人群是7%。B46在象牙海岸人群几乎找不到,在东亚人群是10%。象牙海岸人群主要的B型是B53,在东亚人群里一个没有。这是什么意思?如果病毒变异了,假设它的抗原肽不得不被B53这样的MHC分子呈递,那么东亚人就倒霉了。对于象牙海岸的人来说40%的人是无症状或者很轻,这个病毒很容易控制住。相反,如果一个病毒只能通过B46的MHC分子来呈递它最主要的抗原,东亚人群较象牙海岸人群更好一些。
MHC同种异体多样性是“一箭双雕”。为什么这么说?给定一个MHC分子,你知道它不可能呈递所有抗原肽,一定得有一个可呈递的抗原肽的集合,对应的T细胞受体可以识别这个抗原肽和MHC分子的集合。我们可以用一个灰度谱来表示。在可呈递抗原肽的集合里,一定有一部分是我们自身的抗原肽,它是一个子集。给定一个MHC分子,它就会有类似的自身抗原肽,对应了可以和自身抗原产生反应的T细胞受体的子集。这些T细胞如果天天在血液和淋巴循环里出现就非常危险,可能造成自身免疫病,于是我们的机体产生一种负选择机制,把这样的细胞从我们的T细胞受体集合中删除掉。也就是我们每个人给定一个MHC分子,实际上它潜在可能的TCR集合里有很多缺失。
可以想像,对于给定的一个MHC分子,不论是什么病毒一定会找到缺口,它的抗原只是特定的T细胞受体才可能识别。所以我们种属里如果只有一种MHC分子那就非常危险。
解决的方式是出现多种MHC分子,这是获得性免疫系统非常核心的特点--多样性。不同人之间的MHC分子是不一样的,以至于我们即使有同样自身抗原蛋白的集合,我们每个人身体里的T细胞受体所出现的缺口子集是不一样的,也就是不会有一个病毒因为全人类T细胞的识别不够而彻底的失败,所以历史上很难出现某一种病毒把所有人都消灭掉。历史也证明总是会有人活下来,这种MHC分子的频数在不同地域分布很不同,可能是在古时候的某种病毒或者微生物疾病的爆发导致了某些类型的消失。所以我说它一箭双雕,既解决了自身抗原可能带来的抗自身的免疫反应,同时解决了不是所有人都对某一种病毒产生极其剧烈的敏感性。
有了这样的T细胞反应,有了炎症反应,有了细胞本身固有的免疫机制,我们可以在一定程度上控制大多数的病毒感染。所以可以看到,新冠病毒感染早期到晚期可能有很多无症状感染者或者症状很轻的感染者。我们推测在他的机体内可能发生这样的过程,也就是患者确实被病毒感染了,但是炎症反应和T细胞以及其它获得性免疫细胞的反应在比较早期就控制住了病毒。如果这个过程被拖的很长,炎症反应太大,细胞因子产生太多,病毒复制太厉害,这时候可能机体就无法代偿肺部的呼吸功能,于是出现了重症或者死亡的病例。
我用T细胞的免疫反应给大家讲了固有和获得性免疫以及如何与病毒进行斗争。这里没有提到T细胞反应的局限性,它依赖于MHC的抗原呈递。我刚才说MHC细胞呈递的抗原肽是从病毒来,病毒得先感染一个细胞,才能有RNA翻译产生蛋白。那有什么办法不让病毒跑到细胞里去呢?我们的机体当然有这样的机制,主要就是依靠抗体聚集的过程。抗体分子可以游离于细胞之外,它可以和病毒结合,从而阻断它与细胞受体结合,使病毒被拒之门外的阻断过程我们称之为中和。抗体到底怎么产生呢?这个电影显示病毒被中和的过程。另外抗体还可以产生抵御效应,包被细菌,使它被吞噬细胞吞掉,也就不能起感染的作用。
抗体通过B细胞产生,如上图,紫颜色的是指B细胞产生一种抗体,蓝颜色是产生另外一种抗体,像T细胞一样,每一种B细胞也只产生一种抗体,病毒来了能够识别这种病毒的B细胞识别病毒以后,它就可以被激活。
抗体分子是一个由四个链组成的分子,它有两个轻链、两个重链。轻链和重链的N端都是在不同抗体之间,序列非常不同的地方,我们叫它可变区,这个地方决定了抗体分子和什么样的抗原可以结合,它到底是识别新冠病毒还是识别“旧冠”病毒。它的不变区域,即除了N端可变区之外其他的不变区域,我们叫它恒定区,它决定了抗体分子到底能够产生什么样的免疫应答。为什么机体里能够产生不同的抗体分子, B细胞之所以能够产生不同的抗体,是因为基因组里编码抗体分子重链和轻链的基因区域被拆成很多小片段,如下图所示蓝色绿色和红色。

这样的区域在细胞发生的过程中,也就是在机体骨髓里形成B细胞的过程中,每个B细胞从该区域基因组里挑出一些小片段,组成一个V(D)J的N端的抗体分子,使得每个细胞可以产生不同的抗体。对于某种B细胞来说,它组成的抗体恰恰可以识别某一种抗原比如新冠病毒。那么它就会激活,它激活之后和T细胞一样克隆增殖,有个指数增长,同时它要分化,获得它的效应功能。对于B细胞来说获得效应功能就变成浆细胞,能够大规模产生抗体。抗体释放到血液里,分布到血液里,进入到组织里,可以和病毒结合,通过刚才讲的两种方式,要么阻断了病毒和细胞的吸附,要么把病毒带到某种有清除功能的(比如吞噬功能)细胞里,然后把它消灭掉,而实现免疫保护的作用。
如果仅仅是B细胞产生的抗体,这个简单的过程产生的往往抗体不足以和病毒进行紧密地结合,或者说一开始产生机体免疫反应产生的抗体,亲和力不够高。最后希望能够以最高效率抵抗病毒的抗体,我们把它叫做高亲和力的抗体,它的产生过程是有一个专门的组织结构来完成的,下面看看这个过程到底怎么发生的。
这个过程发生在二级淋巴器官。下图是它的模式图。
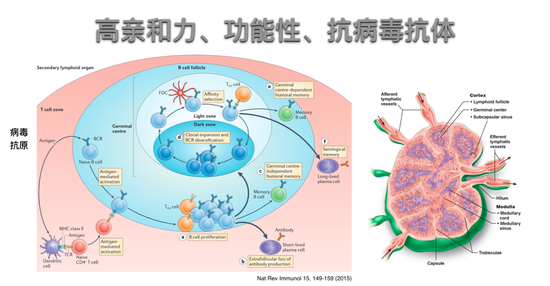
这个模式图显得很乱,有很多种细胞,实际它的原理非常简单和容易理解。试想新冠病毒抗原来了,树突状细胞会把这样的抗原带给T细胞,把T细胞活化,T细胞活化以后有一类T细胞,不是刚刚讲的CD8 T细胞而是CD4 T细胞就到达了B细胞要被活化的地方,等着准备和B细胞进行作用。而B细胞像刚才讲的一样,可以直接识别病毒的抗原,它活化以后就可以直接和T细胞发生作用。
那什么是T细胞和B细胞的相互作用呢?事实上是B细胞一开始它的表面表达着单克隆抗体,既是抗原识别的受体,也是一个内吞受体,它和新冠病毒的抗原结合了以后,就把这个抗原内吞到B细胞内部然后把抗原所产生的肽段呈递到二型MHC分子的表面然后就放到了B细胞表面,这时肽和MHC的复合物就可以被CD4细胞上面的T细胞受体识别,这个T细胞也是被树突状细胞用同样的抗原活化起来,所以这个T细胞和B细胞之间还是抗原特异的相互识别,这个识别使得T细胞就给B细胞传递了一些生化信号,这些信号使得B细胞可以大规模的增殖和分化成抗体生成细胞的过程。
这些信号还有一件非常重要的事情,就是功能性抗体,T细胞产生了很多细胞因子,使得B细胞可以进行抗体的类型转换。什么意思?T细胞产生不同的细胞因子,比如γ干扰素或者IL-4,它就告诉B细胞把抗体的类型换一下,这是什么意思?因为同一个B细胞尽管是克隆性的抗原受体,但是它和抗原结合的V(D)J序列不变,但是它的抗体不变区域可以有好多种不同的类型,依赖于它和T细胞相互作用的收到的细胞因子的信号不同,这个B细胞的后代还是识别同样的抗原,但可产生不同类型的子代。比如,一开始都是产生IgM抗体,后来识别同样的抗原,有的就变成可以产生IgG、IgA的,IgA和IgG对于新冠病毒来说,可能是最重要的,因为IgG效应功能比较强,可以分布到各种各样的组织里。而IgA有一点很独特的地方,它形成了二聚体的时候可以转运到细胞黏膜表面,也就是说呼吸道黏膜表面、消化道黏膜表面,病毒最开始进入机体的时候总是在黏膜表面,如果这里有特异抗体的分子浓度比较高就可以阻断病毒入侵。所以这也是理想情况进行疫苗设计,产生足够高的IgA抗体可能更有效果。
B细胞和T细胞相互作用,B细胞收到信号,进行类型转换同时开始增殖,接下来就进入生发中心,在这里B细胞又干了另一件很神奇的事情,它不停地增殖,同时它还要在它和抗原结合的抗体分子部位得DNA区域引入随机突变。
大家可以想一下,如果在编码区引入随机突变肯定导致氨基酸序列改变,使得这个抗体分子和原来抗原结合的紧密程度不一样。这个发生过程又是B细胞进行增殖的过程,那可以想象这个B细胞产生的子代可能获得这个抗体分子和原来抗原结合的紧密程度就不一样了。于是乎潜在可能出现那些结合力高以及结合力低的抗体分子,机体需要一种筛选的方式把产生结合力高的抗体分子的B细胞筛选下来,让它产生抗体。这个过程在生发中心里由T细胞实现。实现得过程和之前差不多,不同的B细胞的抗体分子,和原来抗原结合的紧密程度不一样,于是获取这个抗原,把它再呈递给T细胞的能力就不一样了。
实际上这些B细胞都在和T细胞进行抗原特异的相互作用。T细胞知道哪个B细胞给他呈递的抗原多,就传递给它更多的信号,而那些收到更多信号的B细胞,最终会变成特别有能力产生抗体,以及长寿浆细胞和记忆B细胞。只有在竞争过程中亲和力比较高的细胞才更又可能成为记忆B细胞和长寿浆细胞。这就是机体如何能力产生出来高亲和力,而且持久的产生抗体的细胞免疫学的过程。当然我们希望新冠病毒感染的时候我们机体的生发中心的反应,输出的就是那些能活下去活的很长的、高亲和力的,可以中和新冠病毒的抗体。
当然,是不是能够中和一个抗原,这件事不是一个免疫系统细胞反应能决定的,这是随机的过程。新冠病毒毕竟太新了,我们所知道的知识相当比较少。
一个抗原,千百个不同抗体
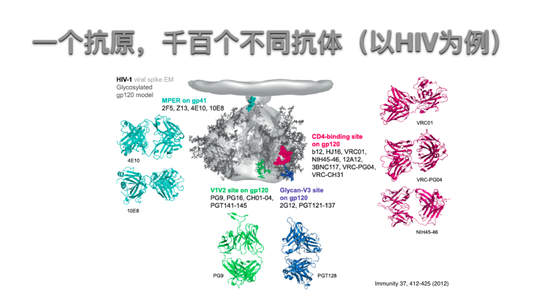
以HIV为例,可以看到一个一个HIV的gp120的抗原,可以有千百个不同的抗体,这些抗体都可以和抗原之间非常紧密的结合,都经历过生发中心的筛选,但是这里真正有用或者真正有中和能力的就是这些有颜色的结合部位的抗体,蓝色、绿色、红色,只有那个抗体和这个部位结合才能阻断病毒和它被感染细胞的黏附。在新冠病毒上相信也是同样,只有一少部分的抗体是有中和活性的抗体。免疫系统本身不可能知道哪个抗体有中和活性,只可能知道哪个抗体可能让B细胞和抗原结合的更紧密。这一点实际上是我们用头脑需要帮助免疫系统,至少让它产生中和抗体更快更多更持久,这就是疫苗的意义。
通过抗病毒疫苗,原则上既可以制造引入T细胞或者CD8 T细胞、杀伤T细胞作用的疫苗,也可以试图促进中和抗体反应的疫苗,或者是两个兼顾。从刚才描述的抗体怎么阻断病毒以及CD8如何杀伤感染细胞的角度,显然最理想是有中和抗体疫苗。
能不能产生中和抗体呢?这个领域现在发展太快,我现在只引用两篇,一篇是清华大学张林琦老师的工作,在恢复期病人的身体里去分离可能产生中和抗体的B细胞。另外一个已经发表的工作是第三军医大学的叶丽林老师发表的。令人鼓舞的是,这些有限的、已经发表的结果表明,康复病人血中确实可以分离到能够特异结合RBD中和抗体的B细胞。这只是说我们能够分离到这种B细胞,并不意味着我们机体里有非常多这样的B细胞,这还是要将来去看,至少有可能我们可以产生中和抗体的疫苗。

谈到疫苗,在我们描述现在疫苗的各种不同路线之前,有必要回顾一下人类疫苗发展的历史的里程碑。大家耳熟能详的肯定是最开始人类与天花搏斗的过程,花了几百年的时间,从中国出现的人痘法,以及外国的“牛痘”法,到1979年彻底消灭掉天花。
这个过程显然我们没有太多免疫学的知识,事实上因为有了这样的实践才使得我们开始问问题,为什么我们机体能够产生这样的免疫反应,才有了今天免疫学。
如果按照现代免疫学概念的建立,上世纪60年代之后发现了B细胞、T细胞,建立了克隆选择学说,才有了现在的免疫学。这之前我们消灭天花的方法已经实现了。而且,另外值得说的是50年代已经有方法可以抵抗脊髓灰质炎,犹太裔的美国科学家SALK,他在50年代的时候开始用细胞来培养病毒的方法,他针对脊髓灰质炎开发了灭活疫苗是有效的。
同时代,俄罗斯裔科学家SABIN开发了减毒脊髓灰质炎活疫苗,我国著名疫苗学家顾方舟老先生从苏联引了种,在我国用了减毒的脊髓灰质炎疫苗,到现在基本上可能除了阿富汗一带,世界上别的地方脊髓灰质炎都已经消灭了。有意思的是在这个发生的过程当中,当然也没有什么现代意义的免疫学知识。那时候由于脊髓灰质炎每年的爆发,不管是中文还是英文世界大家都在问是不是今年就有疫苗呢?我觉得这跟我们今天在问,是不是今年底我们就有新冠的疫苗有非常类似之处。
在有现代免疫学之前,影响人类最重要的两大传染病,在不知道B细胞、T细胞之前就已经可以控制住了。我想说的是,我们的免疫系统其实是很强大的,屁股上、胳膊上扎一针,它还真是可以管用的,是可以产生免疫反应,可以控制病毒,所以对于没有学生物的朋友们不要太焦虑。我也想强调一点,就是有了现代免疫学知识,我们确实可以对非常棘手的一些病原,制造非常有效的病毒。
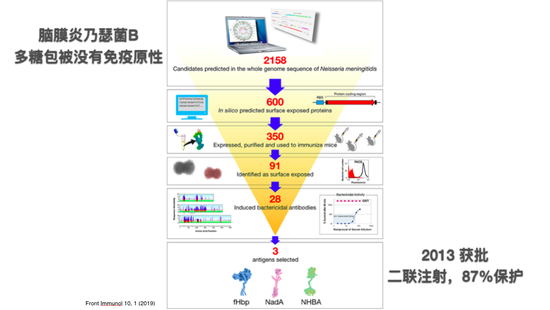
简单举个例子,脑膜炎乃瑟菌B,确确实实用现代免疫学的知识才实现的有效疫苗,因为这个细菌非常奇怪,它不像其他的脑膜炎球菌多糖是有免疫性的,它是打不出来对它多糖的抗体。人们后来发现它的多糖包被和人胚胎发育当中脑子里面的一种脂质非常相似,所以可能机体里能够产生这种抗体的B细胞都被抑制删除掉了,就不产生这样的抗体。后来用反向疫苗学的方法,把这个细菌的全基因组测了序,在里面预测出来有多少可能在表面上所表达的蛋白,差不多600个,把每个单独克隆出来,纯化出来,接种在小鼠体内,然后分离它的血清,看哪一个能够产生抗体,可以在细菌的表面暴露,然后再看能够和这些抗原结合的抗体,哪些可以杀灭病菌的。最后筛选到3到4个抗原,形成复合的疫苗,通过两次注射,确实可以在人体里实现87%的保护,已经很理想了,在2013年获批。这实际上是非常成功的通过现代免疫学的知识,对免疫系统的理解而实现的疫苗保护。这个过程可以看到它还是很漫长,很费劲的。
我们可以像结束天花、结束脊髓灰质炎那样简单易行的方式,还是要经过这样复杂的方式,才能获得新冠疫苗的结果呢,我觉得这个答案显然我们还不知道。
八类疫苗
我们现在已经有的疫苗努力方向有哪些?截至4月30日Nature上总结的全世界范围的疫苗工作大概一百多个研究,大致可以归为八类。

一类,是最古老的减毒活疫苗,现在工作的人相对比较少,它的基本原理就是像天花和脊髓灰质炎一样,用自然方法或者诱导方式使得病毒产生某些突变或者利用一种长的很像但不是同样的病毒,比如牛痘或者马痘的抗原,这样使得免疫系统可以识别抗原,但是我们的机体不会得很严重的疾病,从而实现免疫保护。
另一类是灭活全疫苗,中国的中科生物已经做完一期临床实验,用化学或者物理的方式把病毒彻底灭活掉,不会再产生复制,不再产生疾病了,但是有它的抗原信息,还可以被树突状细胞识别,还可以激活B细胞,从而使得人产生免疫反应。离开了原来的病毒,可以把新冠病毒里面的抗原信息用活病毒载体来表达,然后打到机体里,使得这样的活病毒在细胞里进行复制,产生抗原,提呈给免疫系统,产生免疫反应。也可以用不可复制的载体来表达新冠病毒的抗原,使得机体产生这样的反应。这个过程还是要经历一次表达的过程。

还可以直接把新冠病毒的抗原编码序列以DNA形式或者mRNA形式注入,以DNA形式需要基因枪,DNA需要进入细胞,到细胞核里面,然后转录出来mRNA然后翻译成蛋白,这样的蛋白再提呈到免疫系统里产生免疫反应,或者mRNA直接进入细胞,在胞浆里直接翻译病毒蛋白,产生免疫反应。最后还有直接把抗原给机体注射,亚单位蛋白疫苗,在体外重组出来新冠病毒的S蛋白然后注射到机体里,树突状细胞就会去吞噬,B细胞会去识别,进而产生T细胞或者B细胞免疫反应。
还可以用类病毒颗粒,新冠病毒的蛋白被表达像病毒颗粒的表面,这样的结构,易于刺激免疫系统,使得它认为这是一个外来的潜在的像病毒一样的东西,产生更强的免疫反应,但是它里面没有遗传物质,所以叫类病毒颗粒。
这些不同的疫苗路线,有哪些优劣呢?我觉得显然现在100多种世界各地的实验在尝试所有不同的方向,潜在的可以大致把它们总结,根据他们免疫反应的不同,总结如下,我们考虑它的相对安全性以及免疫原性,是不是能够产生中和抗体,有没有IgA,它是不是能够分布到黏膜的表面,它有多持久,以及它对CD8的应答刺激作用,如图。

打星号的是指可能现在还没有样的疫苗是真正用类病毒颗粒或者DNA和RNA被临床审批过的,但是是潜在可以进行实验的。其他都是有其他传染病是用的,比如亚单位疫苗,乙肝疫苗就是最明显的例子。
安全性来说减毒活疫苗相对来说最不安全,毕竟是活疫苗,取决于它突变多少,有可能突变回程野毒株,这不是不可能的事情。当然历史至少在脊髓灰质炎病毒和天花上有实践,实践上是可行的,只有通过实验才能够知道。免疫原性是说到底产生多强的免疫反应,世界永远都是这样,可能最不安全的也是效果最强的。
减毒活疫苗,如果看已经能够在历史上的疫苗,减毒活病毒所产生的免疫反应总是最强。亚单位疫苗,它的免疫原性相对而言比活病毒差得多。针对新冠病毒来说,现在除了灭活病毒以及减毒活病毒疫苗,它是整个病毒打到体内,其他都还在直接针对S蛋白,主要是因为S蛋白是吸附蛋白,产生的中和抗体可能能够去阻断这样的免疫反应。所以从是不是能够产生中和抗体。原则上所有的疫苗路线都可以产生中和抗体,当然产生中和抗体的滴度的高低,特别是有多持久,可能越是活病毒越可能产生更持久的免疫反应。
针对IgA来说,从它对于保护黏膜更重要这一点来说,减毒活疫苗如果取决于它的给药途径,它很可能可以产生比较高浓度的IgA,但是也不一定,其他的我不知道临床试验将来他们做免疫原形测试的时候,IgA是不是一个终点。CD8的应答,免疫反应过程中它是非常重要的一部分免疫反应,如果仅仅有抗体,你必须在比较早期进行阻断,如果开始有了病毒感染,还是需要有CD8应答。一个完善的疫苗如果能加入CD8的部分,当然是最理想。减毒活疫苗,它的效果往往潜在的可能会更好,但一部分原因是它肯定会激活CD8应答。而亚单位疫苗可能激活CD8应答的能力就会差一些。
预测哪个是最有可能成功的,最有效的,现在为时尚早。发表文献的中可以看到追踪免疫反应,追踪综合抗体,或者抗体免疫反应也就是到三个星期、一个月的情况。康复的病人,他的病毒载量会降低,会产生IgM型的抗体,类型会转化成IgG,会出现中和抗体。但也有研究显示,虽然能产生中和抗体,但中和抗体滴度并不高,这可能需要更广泛的病人研究才能知道。他之所以康复是不是因为这个中和抗体也没有完全定论。最有可能中和抗体是帮助他康复的重要因素,这样的中和抗体到底能够有多持久现在也不明确。
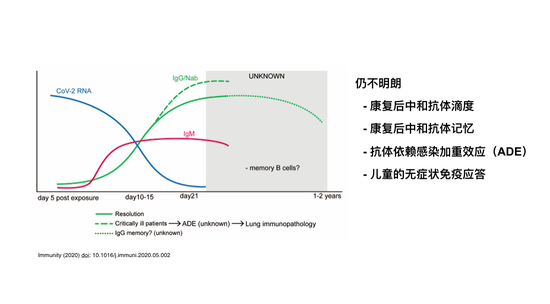
前些日子大家讨论比较多的抗体依赖的感染加重效应或者ADE,在其他冠状病毒里显然是存在的。在新冠病毒是怎么样呢?现在也是悬而未决的需要进一步研究的问题。原则上说,即使是中和抗体也可以产生ADE效应。新冠病毒还有一点,免疫学角度来说,特别有意思的一点,可能告诉我们很重要的信息,就是儿童的无症状免疫应答,好像儿童的无症状比例特别高,儿童产生的免疫反应有什么特殊性,或者特别有效的免疫反应,还是有一种免疫反应在成年、老年之后是有病理作用的而儿童当中没有,这是一个非常重要的悬而未决的要尽快的弄清楚。
有疫苗之前,我们试图用一些治疗方法,也有很多临床试验,我就不详述了。刚才我们讲从早期到晚期,从轻症到重症,从可代偿到失代偿的情况下,里面有一个重要的因素是炎症反应的强烈程度,不停有病毒在复制,不停的有病毒感染,细胞在死亡,不停有炎症细胞产生细胞因子的时候,所以我们叫细胞因子风暴,使得整个体系平衡被打破到一种无法挽回的地步。
对于病人整体来说,尽管很多病毒可能将来你的免疫反应还可以把它清除,但是你已经活不下去了。也有重要的临床实验,在试图用免疫干预的措施,比如托珠单抗来抑制、打断不停地滚雪球的炎症反应,试图能够维持这个病人有足够长时间,允许更健康免疫反应能够清除病毒,把失代偿过程拽回到可缓解的免疫反应过程当中,也是目前很重要的一种尝试。用恢复期病人血清进行治疗,现在临床试验,也是由于我们毕竟只是几个月的时间,下结论还为时尚早。但的确已经有很多基于能够用血清治疗,可能有治疗效果的理论,在开发单克隆抗体以及其他阻断病毒和细胞进行相互作用的治疗方式。
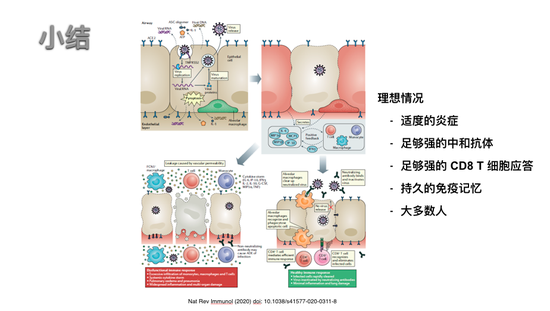
理想情况下我们产生的反应最好是适度的炎症,然后过程中可以产生足够强的中和抗体,足够强的CD8的T细胞应答,不是只是某一种,我们能够恢复从一个炎症反应,回到可缓解,不会失代偿的进程,然后形成持久的免疫。最理想情况是大多数人都是这样,如果某个人实现不了,希望通过疫苗手段帮助每个人的免疫系统达到这种情况,这大致就是我想和大家享的。

当我回顾历史上那些大的瘟疫发生的时候,和今天还有一个非常不一样的地方,除了科学知识不一样,是信息流通的不一样。今天我们处于一种信息爆炸,每一天都在听到有多少人又感染了,有多少人去世了,或者这样那样的问题,使得我们处于持久的焦虑状态。可能对于不直接研究免疫反应的朋友,尤其是感到无助。头绪太多,每个人都在问各种各样的问题。一定程度上这样的好奇是有好处,如果变得非常焦虑的话,从研究免疫角度,持久的焦虑是免疫抑制,可能使你对新冠病毒的潜在易染性更高。
(注:本文根据祁海教授在《理解未来》科学讲座:病毒与人类健康-专题科普”第十一期演讲整理而来,有删减,PPT图片由讲者提供,未经允许不得使用)